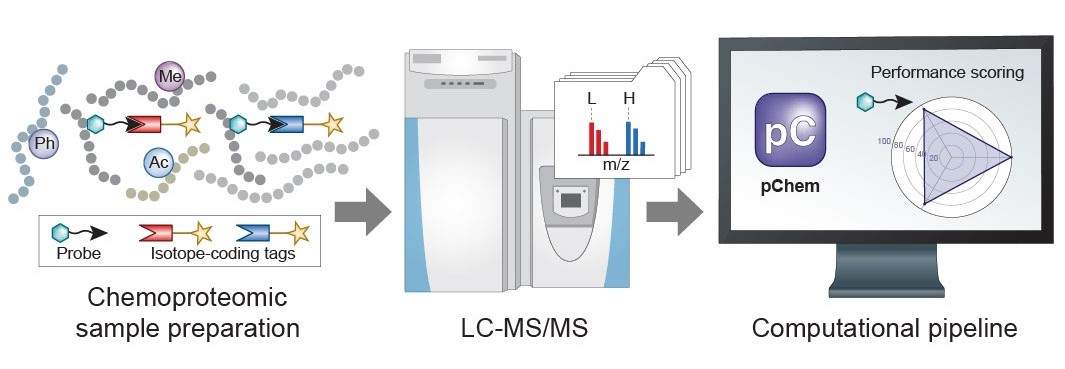
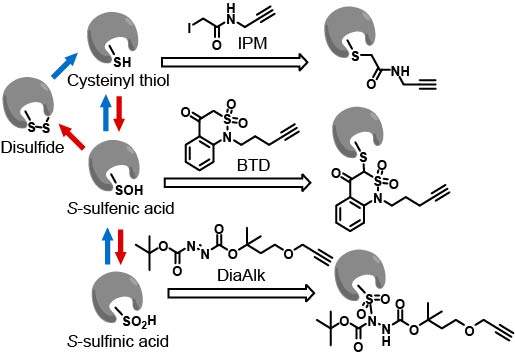
Profiling of the Cysteine Redoxome by Chemoproteomics
Cysteine redox modifications are well-controlled, site-specific cellular events, which play important roles in regulating many biological processes. We have unbiasedly evaluated many redox form-specific chemistries with pChem, enabling
the development of a series of chemoproteomic methods to directly,
site-specifically map and quantify cysteine redoxforms, including cysteinyl
thiol
(-SH, Nat
Protoc 2020)
, sulfenic acid (-SOH, sulfenylation, Nat
Chem 2021), sulfinic acid (-SO2H, sulfinylation
Nat Chem Biol 2018),
and persulfide (-SSH, persulfidation or sulfhydration, Antioxid
Redox Signal 2020).
Our platform has been successfully applied to multicellular animals, such as plants
(Nat Plants, 2022; Proc Natl Acad Sci U S A 2019), worms
(Nat Commun 2021),
flies (Proc Natl Acad Sci U S A 2018),
mice (Nat
Cell Biol 2019) and rats (Redox Biol 2021) for identifying proteome-wide alterations in reactive
cysteines upon oxidative stress or genetically regulated redox perturbation,
providing several valuable resources for uncovering the mechanisms of
redox-modulated control in these model organisms. These data sets not only
provide mechanistic support for prioritizing functional redox sites, but also
offer novel redox mechanisms.